
Who We Are and How We Got Here: Pre-Vertebrate Edition
Prolegomena to any future psychology.


Just as the scientific understanding of human origins has dramatically improved in recent years, so too has the scientific understanding of other branches of the tree of life. I think the pieces are now in place to sketch out the story of how single-celled creatures ultimately gave rise to the complexity of the human mind. Step 1: going from the first cell to something like a lamprey or hagfish.
Farming Fart-Eaters
About 4.5 billion years ago, not long after a protoplanet smashed into the proto-Earth (leading to the formation of the Moon)1, our earliest ancestors — the first cells, or at least the first cells with surviving descendants — came into being. They quickly split into two great families: bacteria and archaeans. By ~1.6 billion years ago2, one group of archaeans had evolved into eukaryotes, whose cells possessed true nuclei3 and, maybe, mitochondria.
About 1.5 billion years ago4, one group of eukaryotes — the archaeplastids — turned cyanobacteria into their domestic servants and thereby gained the ability to photosynthesize. (Present-day archaeplastids include red algae, aquatic green algae, and land plants.) But other eukaryotes had to make a living in a less dignified way: eating bacteria. This is what our ocean-dwelling forebears were up to ~890 million years ago5 when they split off from their closest relatives, the ancestors of present-day choanoflagellates.
Choanoflagellates are unicellular organisms, but several species can also form multicellular colonies that exhibit collective behaviors, like contracting in response to darkness6. It turns out that multicellularity isn’t as special as one might guess: it has emerged independently many times, including in bacteria7. Complex multicellularity, involving differentiated, specialized cell types, seems to have evolved only in eukaryotes, but even that has happened at least five separate times (among red algae, green algae, brown algae, fungi, and — the case at hand — animals)8. So when a group of choanoflagellate-like single-celled organisms began to create larger and more complicated colonies, it wasn’t initially that unusual.
What was the first complexly multicellular animal like? For now, we can only speculate, but here’s my guess: it resembled the bulbous “holdfast” at the bottom of extinct Ediacaran-period (pre-Cambrian) frondose organisms like Akrophyllas:

These Ediacaran biota have long bedeviled paleontologists because they’re hard to classify. There seems to be a soft consensus that they’re more closely related to animals than to any other major clade9, but they don’t look very animal-y. A recent line of research10 suggests that their eye-catching, fractal fronds served to extract oxygen from the water — much like the branching bronchioles of our lungs. But that leaves open the question of how they fed themselves, especially in the deep, dark seafloor areas from which their earliest known fossils originated11.
Following the work of Greg Burzynski12, I think the real action was not in the frond (which may have been a later development13) but in the holdfast that made contact with the sediment. According to Burzynski’s reconstruction, it was hollow, with elaborate interior lobes:

Burzynski has speculated that the holdfast served as something like a bioreactor, providing a favorable environment for the growth of symbiotic bacteria14 that extracted energy from hydrogen sulfide (aka sewer gas or “stink damp”) and methane:

So the bacteria consumed flatulence-like gases, and the ur-animal cultivated the bacteria and periodically ate them15; hence, it farmed fart-eaters.
You’ve Got a Real Smart Mouth
Phylogenetic terminology distinguishes between the concepts of “crown group” and “total group” members of a clade. “Crown group” means, roughly speaking, cousins + their parents + their grandparents; “total group” additionally includes any childless aunts and uncles along the way. I think that the multicellular, bacteria-farming holdfasts were total-group animals, not crown-group animals— lovable spinsters, but not our direct ancestors. Around 830 million years ago16, though, our direct ancestors split off from the holdfasts and began to evolve into something different.
An interesting analogy is carnivory in plants. Carnivorous plants have evolved independently at least five times17 as an adaptation to nutrient-poor habitats. The “snap trap” of the Venus flytrap is especially intricate, with specialized “trigger hairs,” two of which need to be activated over a short time period to close the trap. The decision to close is disseminated in part via action potentials, the same electrical signals used by animal nervous systems18. There’s even a mutant Venus flytrap (called “DYSCALCULIA”) that doesn’t function properly because it fails to integrate these signals: it can’t count19. Carnivorous plants thus offer an existence proof of how natural selection can turn an initially autarkic organism into a (facultative) heterotroph, with sophisticated motor control (as well as extracellular digestion).
We can imagine how something similar could have happened to a holdfast-like creature. Living in a nutrient-poor environment, it developed an upward-facing opening — a mouth — which served to collect bits of detritus — perhaps dead algae drifting down from the photic zone.20 Like a Venus flytrap, this proto–crown-group animal developed ways of detecting when food particles had entered its “trap”, then contracting to create an enclosed cavity to digest the material. Much of the intercellular signaling underlying this system could work fine using broadly diffused molecules like neuropeptides, but, to avoid letting food particles slip away, the management of the mouth had to be faster and more precise. So I think there’s a deep connection between the emergence of the mouth and the advent of the first synapse-forming neurons. Intriguingly, the neurons in ctenophores that most closely resemble on a molecular level the neurons of better-understood animals are those in the mouth region21. The first crown-group animals may have looked like a stripped-down version of the later (but now extinct) ctenophore Xianguangia sinica: a modest (1.5-inch-tall) benthic cup living alongside its bacteria-farming relatives, endowed with a genuine, albeit very basic, nervous system.

Breaking Up the Crown

Around 740 million years ago22, the animal crown group split into two lineages: ctenophores and myriazoans23. I’d guess that this was a case of allopatric speciation: around the same time, at least according to some models, the land mass of present-day South China, where several of the earliest known ctenophore fossils have been found, broke off from the supercontinent Rodinia.24 The proto-ctenophore group, separated from its kin, evolved in its own weird and fascinating direction, ultimately developing a unique version of the nervous system (including a large-scale syncitial nerve net25) and a unique method of locomotion (using “comb plates” each built out of tens of thousands of cilia26).
Meanwhile, the myriazoan lineage changed — evolving a distinct larval life stage that ctenophores lack27 — and diversified. Tempting as it may be to believe that neurons constituted some kind of secret weapon in the evolutionary arms race, it appears that one of the first things that some animals did after getting them was to get rid of them. This was the case for poriferans (sponges), the first major group to branch off after ctenophores. From a distance, early sponges looked similar to the early ctenophores: little cups sitting on the bottom of the ocean. But they developed a different way of life: instead of waiting for bits of algae to show up near their mouths, they actively pumped in water through their porous exteriors, filtering out tiny morsels like bacteria to phagocytose. In this configuration, the ancestral animal mouth became something like a chimney, venting waste water. Fast, precise control over its opening and closing was no longer necessary, so, over time, sponges lost their neurons28.
But the non-sponge myriazoans, called parahoxozoans, retained the old suspension-feeding lifestyle and the basic nervous system that went along with it. They developed more elaborate neuronal control over larger parts of their bodies, using new neurotransmitters like acetylcholine (which explains why nicotine, an acetylcholine-receptor agonist, can induce tentacle contractions29 and maybe peristalsis30 in sea anemones).
Animals managed to scratch out a living during the long “Snowball Earth” glaciations of the Cryogenian period, perhaps benefiting from relatively stable temperatures in the deep ocean31. It was around this time, ~710 million years ago32, that the parahoxozoan group split. One lineage, initially still quite similar to the first crown-group animals, gave rise to present-day cnidarians (sea anemones, corals, jellyfish…) and placozoans. The other lineage ultimately became something very different: bilaterians — motile worms like us.
Not a Cinch
How did a little33 proto-worm emerge from something like a sea anemone? Evidently, it wasn’t easy. One analysis34 identified 157 gene groups that arose in the bilaterian stem lineage. Bilaterians share a whole host of complex, novel adaptations, including the heart, a “through gut” with two openings, a third embryonic germ layer, specific forms of smooth and striated muscle35, primitive eyes of some kind, and the excretory system36. Instead of having neurons scattered throughout their skin (a basiepithelial nerve net), bilaterians developed a segregated, rectangular region of neural tissue situated along the ventral midline — the ur–central nervous system (CNS). Bilaterians’ neurons, unlike their ancestors’, had distinct dendrites and axons (as opposed to generic “processes”), and, for the first time, some of them used monoamine neurotransmitters like serotonin and dopamine37.
How did these profound changes happen? I have a guess. Above, I mentioned in passing that ancestral myriazoans (non-ctenophore crown-group animals) developed a larval life stage. These larvae, which moved around using cilia, had to be at least a little bit smart, analyzing environmental conditions to decide when it was the right time and place to pull the trigger, settle down on the seafloor, and metamorphose into their adult form. In particular, larvae detected and paid attention to various forms of light38. Because larvae were small (and lacked mouths!), they could get by without true neurons, using synapse-less neurosecretory signaling. Meanwhile, their adult forms also had to be at least a little bit smart, but in a different way, using true neurons to control the movement of their mouths and trunks in order to feed. During metamorphosis, the highly light-sensitive, “family planning” intelligence concentrated in the anterior part of the larva disintegrated39, and the more touch-sensitive, feeding-related intelligence concentrated around the mouth took over.
But what if, through a handful of developmental oopsies, the larval intelligence system didn’t go away, and an ectopic adult mouth formed in its vicinity40?
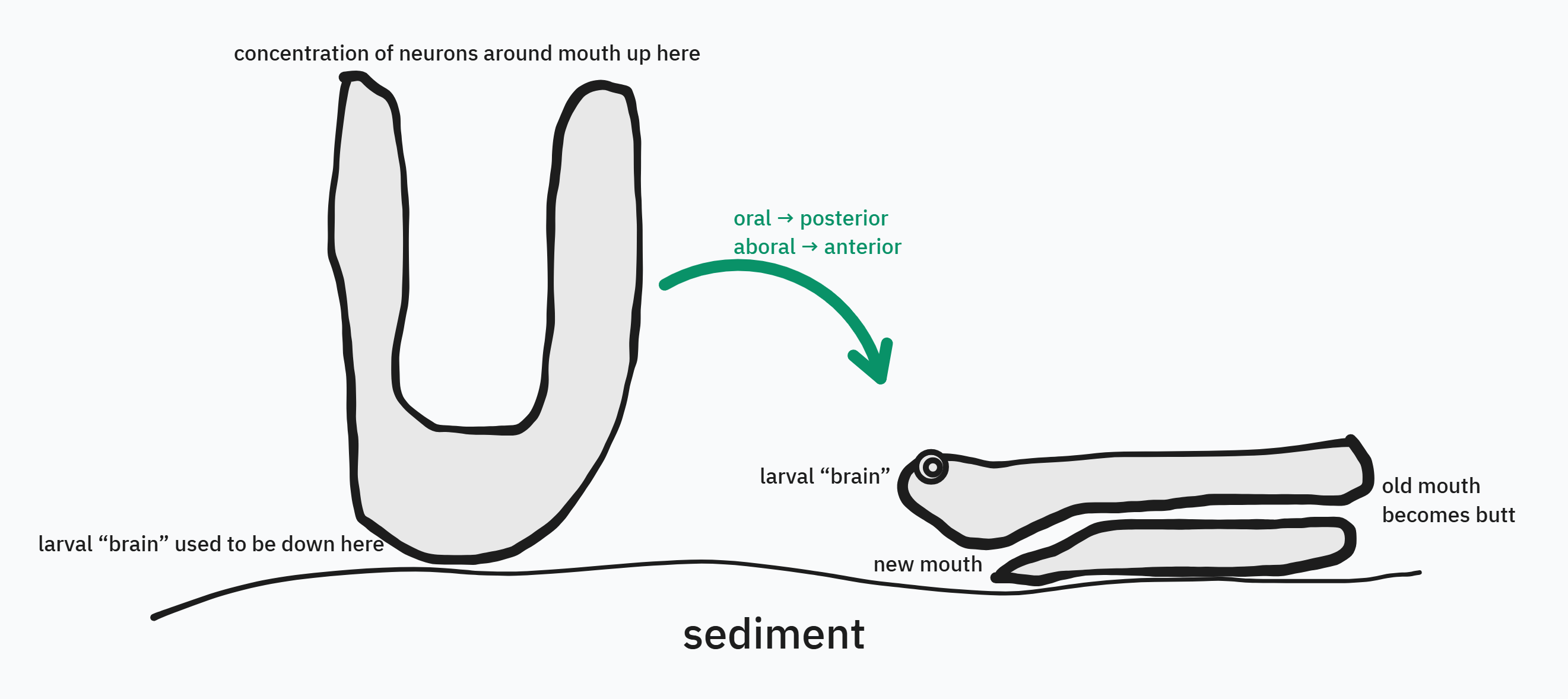
At the end of this transformation, not only was the animal’s body “vermiform” — worm-shaped — but it had, for the first time, a bona fide head. Even at this early stage, its CNS had two parts, which I will call CNShead41 and CNStrunk, and CNShead in turn had two parts: an anterior, light-detecting, “family planning” (metamorphosis- and reproduction-focused) neurosecretory part (homologous to much of the vertebrate hypothalamus42) and a posterior, feeding- and motor-control–focused part,43 which interfaced with CNStrunk to regulate locomotion. Many aspects of this basic nervous-system design are highly conserved across present-day bilaterians, so (as I will hopefully discuss in further detail one day) a lot of claims about the weirdness and incomprehensibility of, say, insect or octopus cognition are probably off-base: we have much more in common than it might appear. Pace Wittgenstein, if a lion could talk, we could totally understand him.
Despite all the ur-bilaterian’s new capabilities, its lifestyle wasn’t very glamorous. Still residing at the bottom of the deep ocean, it wiggled around among its distant bacteria-farming and suspension-feeding cousins, probably scavenging bits of their corpses when they died44. Close to the onset of the Ediacaran period 635 million years ago45, a bilaterian group split off from its relatives and developed the ability to filter-feed by sucking in water and passing it over ciliated slits on the sides of its throat. Humans descend from this group, the deuterostomes, while things like bugs and mollusks descend from the other bilaterian group, the protostomes. By ~590 million years ago46, the ur-deuterostome had begotten the ur-chordate, a filter-feeder that could swim. (Following a suggestion from Adameyko2023, I’m tempted to think that the characteristic chordate hollow neural tube began as an aid to swimming before being superseded by the notochord.) Not much later, the ur-chordate begat the ur-olfactor, which begat the ur-vertebrate — still a swimming filter-feeder, but now clearly recognizable as a fish. Things about its nervous system changed, of course, but I’m more struck by the similarities to the ancestral bilaterian state than any shared vertebrate innovations.
One thing’s for sure: Gary Larson lied. In an old Far Side strip, God muses that it’s easy to create vermiform organisms:
")
But consider the real-life timeline. The first cell was already in place 4.5 billion years ago, mere geological moments after the planet took shape — but it took the next 3.8 billion years to achieve the first proto-worm. These things were not a cinch!

Nor were bilaterians — or animals more broadly — an overnight success. Scientists have thus far found no fossils documenting the earliest phases of their evolution, not just, I suspect, because of unfavorable conditions for preservation47, but also because their ecological range remained quite restricted for a long time48. It was only in the late Ediacaran and, more famously, early Cambrian period that animals began to make an impact on a global scale. We come from a long line of late bloomers.
Appendix: The Meaning of the Monoamines
What’s with serotonin, dopamine, and friends? My irresponsible speculation: in the pre-bilaterian nervous system, sensory neurons often signaled using neuropeptides, with each neuropeptide roughly symbolizing that some state of affairs now obtained in the world. But this was a clunky system: to be able to recognize a different state of affairs, a new neuropeptide/receptor pair had to evolve, and, to be able to recognize a composite state, downstream cells had to express receptors for multiple neuropeptides. Monoamine neurotransmitters evolved as “generalized” neuropeptides, signaling more abstract states in a more modular way. (Monoamine receptors are phylogenetically closely related to neuropeptide receptors, and, as Thiel…2024 points out, neuropeptides often have aromatic amino acids — the raw materials for monoamine neurotransmitters — at their C-terminal ends.)
So what specific states did the different molecules represent in the ancestral bilaterian? I can hazard some guesses:
Serotonin: the presence of light.
Melatonin, synthesized from serotonin: the absence of light.
Dopamine: the presence of food in the mouth/pharynx (hence the deep role in reinforcement learning). (Note that dysphagia (difficulty swallowing) is common in Parkinson’s disease, which is associated with the death of dopaminergic neurons).
Noradrenaline, synthesized from dopamine: the absence of food in the mouth/pharynx (no need to get ready for digestion, go find something to eat).
Tyramine (lost in vertebrates): food is plentiful, conditions are good, chill out.
Combining Betts…2018 and Bowles…2024.
What for? I like the scenario presented in Baum&Spang2023, which centers the physical separation of transcription and translation.
See e.g. Mizuno…2022. Also, Cavalier-Smith2017: “Evolving multicellularity is mechanistically extremely simple. Every unicell group has a cellular and mutational potential to do so given a selective advantage.”
See Burzynski…2020.
Including his dissertation (Burzynski2017).
Burzynski2017: “Alternatively, it may imply that the disc was the ancestral structure, and that the overlying frondose structure evolved later as an oxygen-collection structure under the low and fluctuating oxygenation conditions that characterized Ediacaran seas.”
McIlroy…2021 also proposed that “[l]arge-bodied Ediacaran macrobionts could have gained nutrients from chemosynthetic, sulfur-oxidizing bacteria that likely proliferated at the sediment-water interface around or underneath their bodies”, but the authors there emphasize the fronds over the holdfast and contend that the fronds reclined on the seafloor, contrary, I think, to the common opinion.
Sogin…2020: “There are a number of ways hosts can gain nutrition from their symbionts. Scientists initially hypothesized that hosts ‘milk’ their symbionts to gain nutrients. Milking involves the direct transfer of organic carbon from the symbiont to the host. But there is only incomplete evidence that this is the main mode of nutritional transfer in chemosynthetic symbioses because carbon transfer is challenging to trace and quantify. Also, hosts need other nutrients besides carbon to meet their nutritional needs. In the majority of chemosymbioses, it appears that hosts directly eat their symbionts.”
Not a precise estimate. Derived from my ChronoLog model, dependent here on dates from Liu…2023 and Anderson…2023.
See e.g. Procko…2021.
Bobrovskiy…2020, emphasis added: “Whatever the causes for the proliferation of planktonic algae, their biomass may have fuelled the radiation of eumetazoan animals by increasing the efficiency of nutrient and energy transfer to higher trophic levels based on larger cells sizes compared with bacterial phytoplankton, and by supplying fast sinking food particles to benthic animal communities at the sea floor.”
Derived from my ChronoLog model, dependent here on dates from Liu…2023 and Anderson…2023.
I think Schultz…2023 marked a decisive victory for the “ctenophore first” hypothesis.
See Jokura…2022.
See Wang…2020 and Edgar…2022, though I think this remains a matter of scientific controversy.
Something similar happened later with placozoans: in a sense, they’re all mouth (see DuBuc…2019), plus they’re miniaturized, so their internal coordination can rely on neuropeptide secretion instead of neurons and synapses.
Derived from my ChronoLog model, dependent here on dates from Liu…2023, Anderson…2023, and Sierra&Gold2024.
The oldest known fossilized bilaterian, Ikaria, was only a few millimeters long (Evans…2020), though admittedly it postdates the last common ancestor of crown Bilateria by ~90 million years or so.
See Brunet…2016.
See e.g. Whalan2023. “Opsins originated early in metazoans, and three major opsin lineages, ciliary (c-opsin), Group-4 (RGR/Go) opsins, and rhabdomeric (r-opsin), were already present in the Cnidaria–Bilateria (i.e., eumeatazoan) ancestor” (De Vivo…2023).
See Zang&Nakanishi2020.
This is my spin on the idea of a merger of an apical nervous system and a blastoporal nervous system, which I first encountered in Cisek2019. See Lebedeva…2021 for the homology of the different body axes in cnidarians and bilaterians.
It would be easier to just call this the brain, but the trouble is that, in vertebrates, the hindbrain is anatomically part of the brain but conceptually part of CNStrunk.
In particular, the hp2 prosomere in Luis Puelles’s prosomeric brain model. Puelles’s output is vast, but you can start with Puelles2024.
Homologous to the union of Puelles’s hp1, diencephalon, and mesencephalon in vertebrates.
See Mussini&Dunn2023.
See Anderson…2023.
I would guess that this is why there are no animal fossils among the Weng’an biota of the early Ediacaran (Sun…2024): the Doushantuo Formation derives from shallow water (see e.g. Zhu&Jian2017), and, in the Ediacaran, animals were still exclusively deep-water.
Li…2016. “Flies devoid of OA (TβhnM18) develop a typical couch-potato syndrome, characterised by increased body fat combined with low physical activity. … Flies devoid of both OA and TA did not show this couch-potato phenotype, indicating that OA and TA act antagonistically on major metabolic traits, and particularly on body fat deposition.”
goddammit a *fantastic* post and now I've got to read your back catalog, efffffff.